ASM
1 / ep
1 /
|
|
ORIGINAL ARTICLE |
|
|
|
Translated and adapted by David Ribera-Nebot
|
|
1.
Introduction |
Antoni
Balagué López
|
|
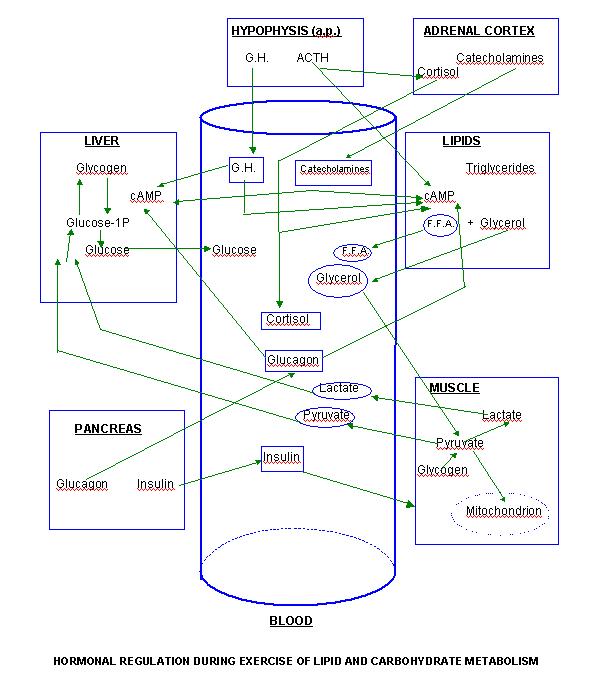
The carbohydrate and lipid metabolic changes that are produced during exercise are induced and regulated by an hormonal substrate. During physical activity, humans develop a complex mechanism of mobilization of selected hormones which regulate and adapt individual’s metabolism to the specific effort situation. This report only deals with the hormones that regulate carbohydrate and lipid metabolism during exercise: catecholamines (adrenaline and noradrenaline), glucagon, insulin, growth hormone and glucocorticoids (cortisol).
|
|
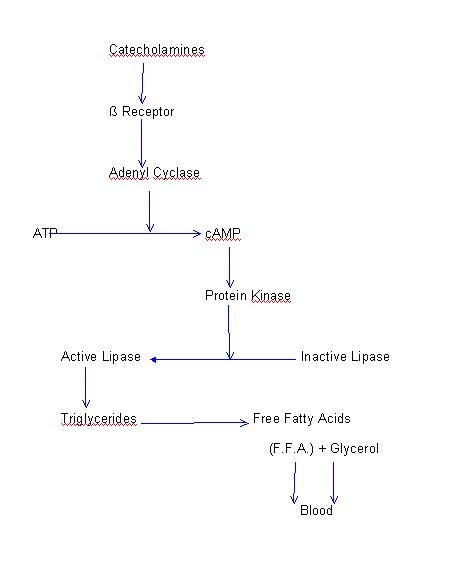
Several authors (Adam, Nowacki and Schmid, 1968; Metze and Linke, 1971; Nowacki, 1969; and Tchibitchian, 1969) have found an increment in urinary excretion of catecholamines or in one of its degradation products, vanilmandelic acid, during exercise. This increment in catecholamines reaches values that are twice the basal ones; however, there is not a significant difference between adrenaline and noradrenaline production (Sarviharjv, 1973). On the other hand, it has been demonstrated that there is a lower urinary excretion of catecholamines in highly trained subjects in comparison with non-trained subjects (Metze, Linke and Mantei, 1971). Regulation During exercise, a sequence of nervous mechanisms determines the increment of the sympathetic autonomic tone. Consequently, nervous stimuli -originated at the motor area of the cerebral cortex- activate sympathetic pathways and, simultaneously, send nervous impulses to the medullar motor neurons. Also, through nervous pathways, the sympathetic pathway is activated by peripheral receptors that are sensitive to the decrease of oxygen concentration (during exercise, muscles uptake considerable amount of O2). Sympathetic autonomic function is reduced with training, and yet there exist no conclusive findings that deeply explain the true mechanism of such adaptation. Metabolic Functions During Exercise a) Effect on the carbohydrates._ Catecholamines activate both hepatic and muscular phosphorylases, and, by doing so, favor glycogen breakdown. The effect of noradrenaline is less than the effect of adrenaline. Specifically, catecholamines favor the synthesis of cAMP , through ATP, which activates the phosphorylase as shown in the following simplified sequence:
|
 |
|
|
|
Company, Balagué and Ballesta (1978) have found a notable increment of cAMP in blood during prolonged swimming; equally relevant, the levels of cAMP remain elevated for 30 minutes after the exercise is finished. These findings corroborate the effect of this nucleotide -cAMP- as a mediator of the glycogenolysis. Through the activation of hepatic phosphorylase the release of glucose into the blood is increased. By means of the activation of muscular phosphorylase, there exists a formation of pyruvate which, in aerobic conditions, is metabolized or, in anaerobic conditions [1], is transformed to lactate that can be oxidized another time in the liver and then favor again the synthesis of glycogen.
[1] In 1980, by trying to clarify a terminology that was not appropriated from a biochemical point of view, Balagué et al. already published a paper (Barbany, Company and Balagué, 1980) pointing out the incorrectness and difficulty to classify the exercise in aerobic and anaerobic. However, this terms are used for practical reasons.
b) Effect on the Lipids._ Catecholamines are able to release fatty acids proceeding from hydrolization of triglycerides that are stored in the fat droplets of lipid cells to the cardiovascular system. Catecholamines activate an inactive lipase converting it into its active form by the cAMP through the protein kinase. The simplified sequence is the following:
|
|
|
|
During effort the increment of the concentration of free fatty acids in plasma may be explained by the increased secretion of catecholamines and cAMP. The difference of free fatty acid concentration in plasma between trained and untrained individuals may be explained, at the same time, by a distinct secretory capacity for catecholamines in both groups. Therefore, in trained subjects the secretion of catecholamines is less and, coincidentally, the increment of fatty acids in blood is also less compared with non-trained subjects (Company, Barbany and Balagué, 1978; Metze, Linke and Mantei, 1971).
|
|
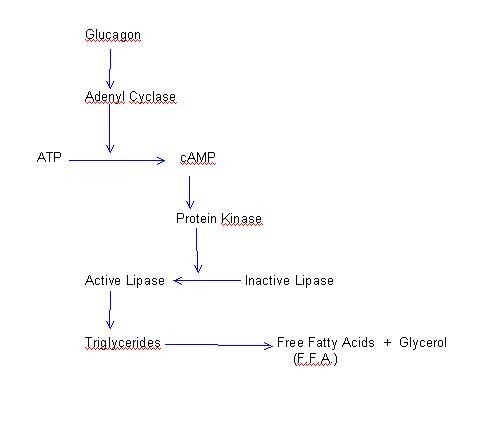
Various authors have demonstrated that the release of glucagon increases during physical exercise (Bottger, Schlein, Faloona Knoche and Unger, 1972; Galbo, Holst and Christensen, 1975; Marliss, Girardier, Seydoux, Wollheim, Kanazawa, Orci, Renold and Porte, 1973; Hellemans, 1978). In exercises of moderate intensity glucagon concentration in blood only increases 35%, whereas in intense and prolonged exercises it may increase as much as 300% (Galbo, Holst and Christensen, 1975). In addition, it have indeed been proved that there exist a completely different response of plasmatic levels of glucagon during exercise between trained and untrained subject. In highly trained subjects there is even a decrement of glucagon levels in blood during exercise and once the exercise activity finishes the glucagon levels return to its normal concentration. On the contrary, in untrained subjects there is a remarkable increase of glucagon levels in blood that remains still elevated two hours after exercise (Hellemans, 1978). Regulation The mechanisms that control the secretion of glucagon during exercise may be classified into two categories: 1. Activators The augment of sympathetic tone produced during exercise, by means of the b-adrenergic pathway, stimulates the secretion of glucagon. For this reason, a beta-blocker (b-adrenergic blocking agent) administered during exercise arrests the increment of glucagon secretion (Marliss, Girardier, Seydoux, Wollheim, Kanazawa, Orci, Renold and Porte, 1973). Some researchers (Galbo, Holst, Christensen, 1975) have found a relationship between the levels of glucagon and the levels of adrenaline and noradrenaline during intense exercise; although this correlation is only significant with adrenaline during moderate exercise. This adrenergic effect is probably mediated by the concentrations of cyclic AMP (cAMP). It have been possible to verify that increasingly concentrations of cAMP injected into an isolated pancreas augment the secretion of glucagon (Weir, Knowton and Martin, 1975). Besides, the great interest in the increments of cAMP concentrations during effort is again emphasized (Company, Balagué and Ballesta, 1978). A decrease of the concentration of blood glucose is a powerful mechanism of secretion of glucagon. However, these decrements of blood glucose concentrations only take place after an intense exercise (Barbany, Balagué and Company, 1978). Consequently, a decrease of blood glucose concentration has significant interest as a secretory stimulus of glucagon only in this situation (intense exercise). An augment of the amino acid concentration in blood bring about an augment of secretion of glucagon. It seems that this effect is not relevant during exercise. 2. Inhibitors The increment of free fatty acids in blood generates a blocking mechanism for the secretion of glucagon. An augment of free fatty acids during exercise was already commented on in the previous chapter (Company, Barbany, and Balagué, 1978). As a result, a negative control by free fatty acids must be conferred depending on an individual’s training state. The increments of glucagon concentration during exercise indicate that there exists a predominance of the activator over the inhibitor effects of glucagon. Metabolic Functions During Exercise a) Effect on Carbohydrates._ Glucagon stimulates glycogenolysis and gluconeogenesis. Glycogenolytic._ Glucagon stimulates the adenincyclase of hepatic cells. This leads to an activation of phosphorylase and, therefore, an increment of glycogen breakdown. Glucagon does not bring about glycogenolysis in the muscle. Glyconeogenesis in the liver increases as a function of the available amino acids. b) Effect on Lipids._ The intracellular lipase of adipose tissue, which transforms triglycerides into free fatty acids and glycerol, is also sensitive to the action of glucagon. The mechanism by which this lipase is activated is similar to the mechanism commented previously:
|
|
|
|
Thus, this lipolytic effect of glucagon also participates in the increment of free fatty acids generated during exercise. In this case, also there exists a parallelism between the degree of training and the blood levels of glucagon and free fatty acids.
|
|
Some authors ( Hoffman, Fiedler and Kibittel, 1972; Pruett, 1970; Gerberger, Keibel, Langer and Pickenhain) demonstrate a decrease of plasmatic concentrations of insulin during effort. However, Hellemans (1978) proves a divergent behavior of plasmatic insulin levels during effort depending on the training state of subjects. In trained individuals there exists a rise in the levels of plasmatic insulin during the first three minutes of effort on a cycle ergometer, then plasmatic insulin decrease progressively and at the end of exercise its levels are lower than the basal levels. Also, two hours after exercise the plasmatic insulin levels remain under the basal levels. In contrast, in untrained individuals the levels of plasmatic insulin increase quickly during the first minute of effort, then these levels remain elevated throughout all the effort and even two hours after the exercise is finished. These findings agree with previous findings (Rottini, Cozzolino, Dominici and D’ovidio, 1971) in which untrained individuals also increased their levels of plasmatic insulin during exercise. Equally important, the concentrations of plasmatic insulin during effort of short duration vary depending on the ingestion of a carbohydrates-rich solution before exercise (Orara, Vapaatalo, Saarela and Reinila, 1974). In individuals who ingest water without carbohydrates (placebo), plasmatic insulin drops during exercise under initial levels; whilst in individuals who ingest carbohydrates, although plasmatic insulin decreases during exercise, it never drops under initial levels. This experience suggests that there exists a modification of the efficacy of the stimuli that are responsible for the variation of secretion of insulin during exercise, based upon the levels of glucose in the blood. Regulation The secretion of pancreatic insulin during effort is regulated by: The Levels of Glucose Concentration in the Blood. This does not seem to be a relevant regulatory mechanism during exercise since the plasmatic levels of glucose vary little. Likewise, when the concentration of blood glucose is low, this regulatory mechanism might generate a blocking stimulus for the secretion of insulin; therefore, it may not be relevant for intense and prolonged exercise either. The Levels of Plasmatic Glucagon. Glucagon is a stimulative hormone of the secretion of insulin by means of a direct action on beta cells and by means of an indirect action through the increments of glucose in the blood. Moreover, it must be pointed out that during exercise the increments of glucagon produce an activator direct effect on the secretion of insulin. The Concentration of Catecholamines in the Blood. The augments of noradrenaline in the blood inhibit the secretion of insulin (Orara, Vapaatalo, Saarela and Reinila, 1974; Karam, Grasso, Wegienka, Frodsky and Forsham, 1966). Accordingly, the increments in secretion of catecholamines that are produced during exercise will function as an important blocking mechanism of the pancreatic secretion of insulin. The Blood Levels of cAMP._ The augments in the blood of cAMP stimulate the secretion of insulin. Previously, it has already been commented that the blood concentration of cAMP increases during exercise. The Somatostatin._ The increments of somatostatin block the secretion of insulin. During exercise there exists an increment of the growth hormone and, very probably, the release of somatostatin is blocked. To summarize, during exercise there exist various activator and inhibitor mechanisms of the secretion of insulin that act simultaneously. This fact helps to explain the findings of different researchers, contradictory in part, about the plasmatic modifications of insulin during exercise, especially when considering the variations of hormonal secretions in relation to the training state of the individual. Metabolic Functions During Exercise a) Effect on the Carbohydrates._ The increase of the plasmatic secretion of insulin produces: Increment of glucose uptake by the muscle and by other tissues through an action on the cellular membrane. Increment of glucose uptake by adipose tissue. Increment of muscle glycogen synthesis. Decrement of cAMP, of gluconeogenesis and of the liver glycogen synthesis. The relevant effect that insulin could produce on the muscular uptake of glucose during exercise remains doubtful for, at least, two reasons: (Company, Barbany y Balagué, 1978) because of the fluctuations (increments or decrements) in the blood levels of plasmatic insulin that are produced during effort; and (Barbany, Balagué y Company, 1978) due to the corroborative fact that the uptake of glucose by skeletal muscles during exercise increases even when insulin is absent (Ganong, 1976). b) Effect on the Lipids._ The increase of the plasmatic secretion of insulin produces: Increment of the synthesis of fatty acids in adipose tissue. Increment of the synthesis of glycerol phosphate in adipose tissue. Increment of the storage of triglycerides in adipose tissue. Increment of the synthesis of lipids in the liver. The doubts about the physiological importance of insulin during exercise have already been commented in the previous paragraph.
|
|
It seems clear that the plasmatic concentrations of growth hormone increase during exercise (Buckler, 1971; Roth, Glick, Yalow and Berson, 1963; Pop, Cioara, Dragan and Vta, 1975; Sutton, Young, Lazarus, Kickie and Markuytis, 1960). An effort test on a cycle ergometer has even been used as a clinic test to explore the hypophysis release of growth hormone (Buckler, 1972). The plasmatic levels of growth hormone during exercise change with training. In well-trained subjects the augments of growth hormone in the blood during exercise are less marked than in non-trained subjects. Furthermore, once exercise is finished, the elevated levels of this hormone continue for a longer period of time in non-trained subjects as opposed to well-trained subjects (Sutton, Young, Lazarus, Kickie and Markuytis, 1960). Regulation The mechanisms responsible for the increment of growth hormone during exercise are not well understood. However, the following points must be considered: Hypoglycemia stimulates the secretion of growth hormone. This does not seem to be the usual responsible mechanism during exercise because of the identical reasons that have been commented previously. Glucagon stimulates the secretion of growth hormone, and during exercise the plasmatic levels of glucagon increase notably. A direct effect of the motor area on the hyphotalamus may exist, governing the secretion of growth hormone through nervous control. High levels of lactate in the blood stimulate the secretion of growth hormone. In fact, when lactate is injected into the venous system, a significant increase of the concentration of plasmatic growth hormone occurs (Sutton, Young, Lazarus, Kickie and Markuytis, 1960). This experience might explain a coordination between the recovery of plasmatic lactate and growth hormone to normal values after exercise, a process that is faster in individuals with a higher degree of training. In other words, it must be emphasized that the higher the level of an individual’s training, the shorter this mentioned recovery. We should ask ourselves if blood lactate levels may be a relevant factor for the secretion of growth hormone during effort. Metabolic Functions During Exercise a) Effect on the Carbohydrates._ During an initial phase of exercise, growth hormone increases the capacity for glucose uptake by muscles. During a more advanced phase of exercise, the muscular capacity for glucose uptake is inhibited and, simultaneously, the secretion of glucose by hepatic glycogen is increased. This hepatic glycogenolytic action is attributed to the effect of growth hormone favoring the synthesis of specific proteins which are necessary for the synthesis of cAMP.(Young, 1968). b) Effect on the Lipids._ During an initial phase of exercise, growth hormone favors muscular uptake of fatty acids (Young, 1968). During an advanced phase of exercise, it favors lipolysis through the synthesis of cAMP. On the whole, growth hormone, together with catecholamines and glucagon, may produce an important effect on the increment of free fatty acids that are released in the blood during exercise.
|
|
The levels cortisol in the blood increase notably during exercise; moreover, the higher the intensity of the effort, the more marked the increments of cortisol. On the other hand, the increments of plasmatic cortisol are lower in trained individuals (Keibel, 1974). The changes of cortisol are originated at central level, varying the levels of ACTH with exercise. A maximal intensity exercise provoke an increment in the secretion of hypophysary ACTH (Fortuno, Fraioli and Isipori). On the other hand, the response of adrenal cortex to a venous administration of ACTH, is the same for both trained and untrained individuals (Frenkl, Csalay, Scakrar and Langfy, 1971). Regulation The clarification of the mechanisms that contribute to increase the secretion of ACTH and glucocorticoids during exercise is a difficult endeavor. One could easily explain these mechanisms based upon the “stress” concept, by conferring a double role on exercise: a stressful role for the organism and a causative role for the secretion of both ACTH and cortisol. In contrast, one should think of a more physiological mechanism; therefore, it cannot be underestimated the capability of the adrenal cortex, concretely by its motor functions, for activating regulatory centers of cardiac activity, vasomotor tone and respiration; consequently, the adrenal cortex might govern the glucocorticoid function through the activation of the hypothalamic secretion of RF-ACTH. Metabolic Functions During Exercise a) Effect on the Carbohydrates._ It is classically known that the administration of glucocorticoids provokes an elevation of blood glucose levels together with an increment in the content of hepatic and muscular glycogen. These effects are due to a glyconeogenic effect which is fundamentally mediated by amino acids, lactate and pyruvate. The glyconeogenic effect that is mediated by lipids is not, apparently, so relevant for the regulation of glucocorticoids. b) Effect on the Lipids._ ACTH possesses a lipolytic effect, activating the adenin cyclase that simultaneously activates the lipase of fat cells. Glucocorticoids also increase the lipase activity, even though it is through a much slower process which requires the synthesis of a new protein and is independent of the effects of cAMP.
|
|
|
|
|
|
|
|
Hormone:
{G. hormòn, pres. part. of hormaõ, to rouse or set in motion}. A
chemical substance, formed in one organ or part of the body and carried
in the blood to another organ or part; depending on the specificity of
their effects, h.’s can alter the functional activity and sometimes
the structure, of just one organ or of various number of them. Catecholamines:
Pyrocatechols with an alkylamine side chain. Examples of biochemical
interest are epinephrine, norepinephrine and dopa. Dopa:
3,4- dihydroxyphenylalanine; an intermediate in the catabolism of
phenylalanine and tyrosine and in the biosynthesis of norepinephrine,
epinephrine and melanin; the L form, levodopa, is biologically active. Epinephrine=Adrenaline:
3,4-dihydroxy-a
{methylaminomethyl} benzyl alcohol; a catecholamine that is the chief
neurohormone of the adrenal medulla of most species. It is the most potent
stimulant (sympathomimetic) of adrenergic a
and b
receptors. Norepinephrine=Noradrenaline:
Levarterenol; 1-a-(aminomethyl)-3-4-dihydroxybenzyl
alcohol; a catecholamine hormone of which the natural form is D,
although the L form has some activity; the base is considered to be the
postganglionic adrenergic mediator. It is present in the adrenal medulla
and in adult animals of most species in much smaller amounts than is
epinephrine; possesses the excitatory actions of epinephrine, but has
minimal inhibitory effects. Glucagon:
Hyperglycemic-glycogenolitic factor; HG factor; pancreatic hyperglycemic
hormone; a hormone consisting of a straight-chain polypeptide of 29
residues (bovine g.) extracted from pancreatic alpha cells. Insulin:
A peptide hormone, secreted by beta cells in the islets of Langerhans,
that promotes glucose utilization, protein synthesis and the formation
and storage of neutral lipids; obtained from various animals and
available in a variety of preparations. Somatostatin:
Somatotropin release-inhibiting factor; a tetradecapeptide capable of
inhibiting the release of somatotropin by the anterior lobe of the
pituitary gland. Growth
Hormone (GH)=Somatotropin: Growth or pituitary growth hormone;
somatotropic hormone; a protein hormone of the anterior lobe of the
pituitary, produced by the acidophil cells, that promotes body growth,
fat mobilization and inhibition of glucose utilization; diabetogenic
when present in excess. Glucocorticoid:
Glycocorticoid. Any steroid-like compound capable of significantly
influencing intermediary metabolism such as promotion of hepatic
glycogen deposition and of exerting a clinically useful
anti-inflammatory effect. Cortisol is the most potent of the naturally
occurring g.’s; most semisynthetic g.’s are cortisol derivates. Cortisol:
Hydrocortisone. Cortisone:
{former acronym for corticosterone}. 17a,
21-dihydroxy-4-pregnene-3, 11,20-trione; 17a-hydroxy-11-dehydrocorticosterone;
a glucocorticoid not normally secreted in significant quantities by the
human adrenal cortex. ACTH=Adrenocorticotropic
Hormone: corticotropin; adrenocorticotropin; adrenotropin;
corticotropic or adrenotropic h.; the h. of the anterior lobe of the
hypophysis which governs the nutrition and growth of the adrenal cortex,
stimulates it to functional activity and also possesses extraadrenal
adipokinetic activity; it is a polypeptide containing 39 aminoacids, but
exact structure varies from one species to another; sometimes prefixed
by a
to distinguish it from b-corticotropin.
|
|
|
|
ADAM, K., NOWACKI, P. und SCHMID, E. (1968). Untersuchugen über die Sympathico-Adrenale Reaktion bei Hochleistungsportlern im Training und im Wettkampf. Sportartz v. Sport Med., 19, 389. BARBANY, J.R., BALAGUÉ, A. y COMPANY, X. (1978). Metabolismo de los Carbohidratos en el Ejercicio. Apuntes de Medicina Deportiva, 60 (15). BOTTGER, I. SCHLEIN, E. FALOONA, G.R., KNOCHEL,J.P. and UNGER,R.H. (1972). The Effect of Exercise on Glucagon Secretion. J. Clin. Endocrinol. Metab., 35, 117. BUCKLER, J. (1971). Growth Hormone Levels with Exercise. Arch. Dis. Child., 46, 399. COMPANY, X., BALAGUÉ, A. y BALLESTA, A. (1978). Algunas Modificaciones de la Mioglobina Sérica en un Grupo de Nadadores Durante el Esfuerzo. Comunicación presentada al I Congreso Nacional de la Sociedad Española de Química Clínica. Barcelona, octubre 1978. COMPANY, X., BARBANY,J.R. y BALAGUÉ, A (1978). Metabolismo Lipídico en el Ejercicio. Apuntes de Medicina Deportiva, 59 (15). BARBANY, J.R., COMPANY, X. y BALAGUÉ, A. (1980). Bases Bioquímicas del Trabajo Muscular Anaeróbico. Apuntes de Medicina Deportiva, 65 (17), 9-14. BUCKLER, J.M.H. (1972). Exercise as a Screening Test for Growth Hormone Release. Acta Endocrinologica, 69, 219. FORTUNO, G., FRAIOLI, F. and ISIPORI, A. Valutazione della Risposta Incretora Ipofisaria all’Activitá Fisica. FRENKL, R., CSALAY, L., SCAKRAR, G. und LANGFY, G. (1971). Untersuchung der ACTH-Wirkung auf den Steroidspiegel des Plasmas im Trainerten und im Untrainierten Organismus. Med. und Sport, 4, 122. GALBO, H., HOLST, J.J. and CHRISTENSEN, W.J. (1975). Glucagon and Plasma Catecholamine Response to Graded and Prolonged Exercise in Man. Journal of Applied Physiology, 38, 70. GANONG, W.F. (1976). Manual de Fisiología Médica. Edt. El Manual Moderno, página 294. GERBERGER, G., KEIBEL, D., LANGER, H. und PICKENHAIN, L. (1975). Steverungssebenen Molekulare Mechanismen und Dynamik der Hormonellen Regulation im Menschlichen rganismus. Med. v. Sport, 15, 97. HELLEMANS, J. (1978). Het Energie Metabolisme bij Getrainde en Ongetrainde individuen. Gennees-Kunde en Sport, 6, 130. HOFFMANN, H.D., FIEDLER, H. und KIBITTEL, W. (1972). Der Einfluss von Fahrradergometerait Sparring und Wettkampt auf die Plasmainsulin Konzentration und den Blutzuckerspiegel bei Boxern. Med. und Sport, 4, 119. KARAM, J.H., GRASSO, S.G., WEGIENKA, L.C., FRODSKY, G.M. and FORSHAM, J.H. (1966). Effect of Selected Hexoses, of Epinephrine and of Glucagon on Insulin Secretion in Man. Diabetes, 15, 571. KEIBEL, D. (1974). Nebemniereninden-Hormone und Sport Liche Leistung. Med. und Sport, 65 (3). MARLISS, E.B., GIRARDIER, L., SEYDOUX, J., WOLLHEIM, C.B., KANAZAWA, Y., ORCI, L., RENOLD, A.E. and PORTE, D. (1973). Glucagon Release Included by Pancreatic Nerve Stimulation in the Dog. J. Clin. Invest., 52, 1246. METZE, R., und LINKE, P.G. (1971). Adrenalin und Noradrenalin Ausscheidung bei Fußballspielern. Mediz. v. Sport, 23 (1). METZE, R., LINKE, P.G. und MANTEI, E. (1971). Der Katekolaminumsatz bei Trainierten und Untrainierten Jugendlichen und Erwachsenen. Med. v. Sport, 11, 327. NOWACKI, P. (1969). The Turnover of Sympathico Adrenal Hormones of Sportsmen in Training Anticipation and During Competition Judged by Measurements of the Urinary Excretion of 3-metnoxy 4-hydroxy Mandelic Acid. Biochemistry of Exercise. Medicine and Sport, 3, 205. Karger: Basel. ORARA, S., VAPAATALO, H., SAARELA, J. and REINILA, M. (1974). Blood Glucose, Serum F.F.A. and Serum Insulin Levels After the Intake of the Carbohydrate Rich Solution Before Exercise in Man. The Journal of Sports Medicine and Physical Fitness, 73 (14) POP, T., CIOARA, Z., DRAGAN, J. and VTA, I. (1975). Recherches Concernant les Variations de l´Hormone de Croissance Plasmatique chez les Esportifs de Haute Competition sans l’Influence au stress Psychique. Medicine du Sport, 38 (1). PORTE, D., GRABER, A., KUZUYA, T. and WILLIAMS, R.H. (1966). The Effect of Epinephrine on Immunoreative Insuline Levels in Man. J. Clin. Invest., 45, 228. PRUET, E.D.R. (1970). Plasma Insulin Concentrations During Prolonged Work at Near Maximal Oxygen Uptake.Journal of Applied Physiology, 29, 155. ROTH, J., GLICK, S.M., YALOW, R.S. and BERSON, S.A. (1963). Secretion of Human Growth Hormone: Physiologyc and Experimental Modification. Metabolism, 12, 577. ROTTINI, E., COZZOLINO, G., DOMINICI, G. and D’OVIDIO, M. (1971). Physical Activity and Blood Insulin, Sugar and NEFA Valves in Normal Untrained Subjects. The Journal of Sports Medicine and Physical Fitness, 1 (1). SARVIHARJV, P.J. (1973). Effect of Physical Exercise on the Urinary Excretion of Catecholamines and 17-hidroxycorticoteroides in Young Healthy Men. The Journal of Sports Medicine and Physical Fitness, 14, 171. SUTON, J.B., YOUNG, J.D., LAZARUS, L., HICKIE, J.B. and MARKUYTIS, J. (1960). The Hormonal Response to Physicl Exercise. Austr. Ann. Med., 84 (18). TCHIBITCHIAN, D.A. (1969). Sostoinanie Simpato-Adrenalovoi Sistemy ou Iounykg Legkoatletov pri Fizitecheskoi Nagrouzke na Vynoslivost. Teor. Prakt. Fiz. Kovlt, 2, (32). YOUNG, F.G. (1986). Growth Hormone. Edit. A, Pecile and E.E. Muller. Excerpta Médica: Amsterdam, pág. 139. WEIR, G.C., KNOWTON, S.D. and MARTIN, D.B. (1975). Nucleotide and Nucleoside Stimulation of Glucagon Secretion. Endocrinology, 97, 932.
|
|
|
| © 1979 Antoni Balagué López ALL RIGHTS RESERVED |
 © 2003 humanmovement.com |